Multiple Functions and Regulation of Mammalian Peroxiredoxins
近期要做的研究内容与 PRDX2 相关,正所谓知己知彼百战不殆,读点相关的综述也是必须的喵!~o( =∩ω∩= )m
点此链接阅读原文。
本次来带的综述讲述了 哺乳动物过氧化物酶 (Peroxiredoxins, Prxs),这是一类高亲和力、对 \(\ce{H2O2}\) 高度敏感的硫醇依赖性过氧化物酶家族。综述提到了几个核心发现:
- 作为抗氧化剂消除毒性过氧化物。
- 在特定亚细胞部位通过 可逆性失活 局部调控 \(\ce{H2O2}\) 信号分子浓度梯度。
- 作为 \(\ce{H2O2}\) 信号的传感器与转换器。
Abstract
过氧化物酶 (Prx) 是一类重要的过氧化物酶,哺乳动物细胞表达六种 Prx 亚型(PrxI 至 PrxVI)。细胞在细胞内不同部位产生过氧化氢 (H₂O₂), H₂O₂ 可作为信号分子发挥作用。鉴于 Prx 含量丰富,且其活性位点上的半胱氨酸 (Cys) 残基对 H₂O₂ 的氧化高度敏感,因此这种氧化剂的信号功能需要广泛且高度局部的调控。本文综述了近期关于 PrxI 在中心体通过磷酸化进行可逆调控以及 PrxIII 在线粒体中活性位点 Cys 过度氧化的研究成果,作为 H₂O₂ 信号局部调控的实例。此外,Prx 对 H₂O₂ 氧化具有很高的亲和力和敏感性,使其能够通过将其氧化态转移给结合的效应蛋白,作为 H₂O₂ 信号的传感器和转换器。
1. Introduction
过氧化物还原酶 (Prx) 是一类 硫醇依赖性过氧化物酶家族,催化 \(\ce{H2O2}\)、烷基氢过氧化物和过氧亚硝酸盐的还原反应。
结构特点
所有 Prx 酶在 N 端区域含有一个保守的 Cys 残基(称为过氧化物酶 Cys, \(\ce{C_P}\)),该残基是过氧化物氧化的位点。
大多数 Prx 酶在 C 端区域含有一个额外的保守 Cys 残基 (还原酶 Cys, \(\ce{C_R}\))。
根据 \(\ce{C_R}\) 是否存在与所在位置,可以将 Prx 分为 2-Cys 亚家族、非典型 2-Cys 亚家族和 1-Cys 亚家族。
哺乳动物表达六种过氧化物酶同工酶:
- PrxI-PrxIV: 2-Cys 亚家族,主要利用 \(\ce{H2O2}\)。
- PrxV: 非典型 2-Cys 亚家族,偏好烷基氢过氧化物与过氧亚硝酸盐。
- PrxVI: 1-Cys 亚家族,偏好烷基氢过氧化物。
Prx 的结构使其 \({\ce{C_P-SH}}\) 具有对过氧化物的高度亲和力,从而承担对抗有毒过氧化物的职责。不过在最近的发现中,Prx 被指出可能还存在更多样的功能:
- 通过在不同的亚细胞区室中发生过氧化物酶活性的可逆性失活来调节局部 \(\ce{H2O2}\) 浓度。
- 可逆性失活: 基于 \({\ce{C_P-SH}}\) 的过度氧化或 Tyr/Thr 磷酸化实现。
- 在类固醇生成、昼夜节律、生长因子信号传导和细胞周期进程中发挥重要的生理作用。
- 反映了 \(\ce{H2O2}\) 作为胞内信使的身份承担重要调控功能。
- Prx 作为 \(\ce{H2O2}\) 信号通路的传感器与转化器发挥作用。
2. Peroxidase Reaction Mechanisms
所有生物的 Prx 酶均为 专性二聚体,并含有一个绝对保守的 N 端 \(\ce{C_{P}-SH}\) 残基,该残基与 \(\ce{H2O2}\) 反应生成 \(\ce{C_P-SOH}\)并释放 \(\ce{H2O}\)。
- 在 2-Cys Prx 中,亚磺酸中间体会与二聚体的另一个 Prx 的 \(\ce{C_R}\) 进攻形成 二硫键。随后该二硫键会被 硫氧还蛋白 (Trx) 还原。
- 2-Cys Prx 二聚体的两个亚基反向平行排列,并通过 \(\ce{Cys^{52}-Cys^{173}}\) 二硫键相连。
- 非典型 2-Cys Prx 与 1-Cys Prx (PrxV/PrxVI) 初始步骤与 2-Cys Prx 相同,但无需另一亚基的协助即可完成亚磺酸的还原。
- PrxV 形成亚基内 \(\ce{Cys^{52}-Cys^{173}}\) 二硫键,并可被 Trx 还原。值得注意的是,PrxV 二聚体为非反向平行排列。
- PrxVI 缺乏 \(\ce{C_R}\),必须依靠 π 型谷胱甘肽 S-转移酶 (πGST) 和两分子的 谷胱甘肽 (GSH) 形成二硫键并被还原。
专性二聚体
在正常的生理条件下,这些蛋白质在物理结构上必须以二聚体(或由二聚体组装成的更高阶寡聚体)的形式存在,它们无法单独作为孤立的单体稳定存在。
对于 Prx 酶来说,它们在细胞内天然两两成对存在,以保证活性位点的结构正确。(即使它们之间没有形成二硫键)

3. Active Site Structure and Exceptional Reactivity of the Peroxidas Cysteine
对于 Cys 是否能胜任过氧化物酶的氧化还原中心这件事,在一开始是饱受质疑的。一方面是因为早期科学界所知的所有过氧化物酶都以金属辅因子或硒代半胱氨酸为活性中心;另一方面,如何在蛋白质众多 Cys 残基中选择性靶向充当活性中心的 Cys 残基也是未解之谜。
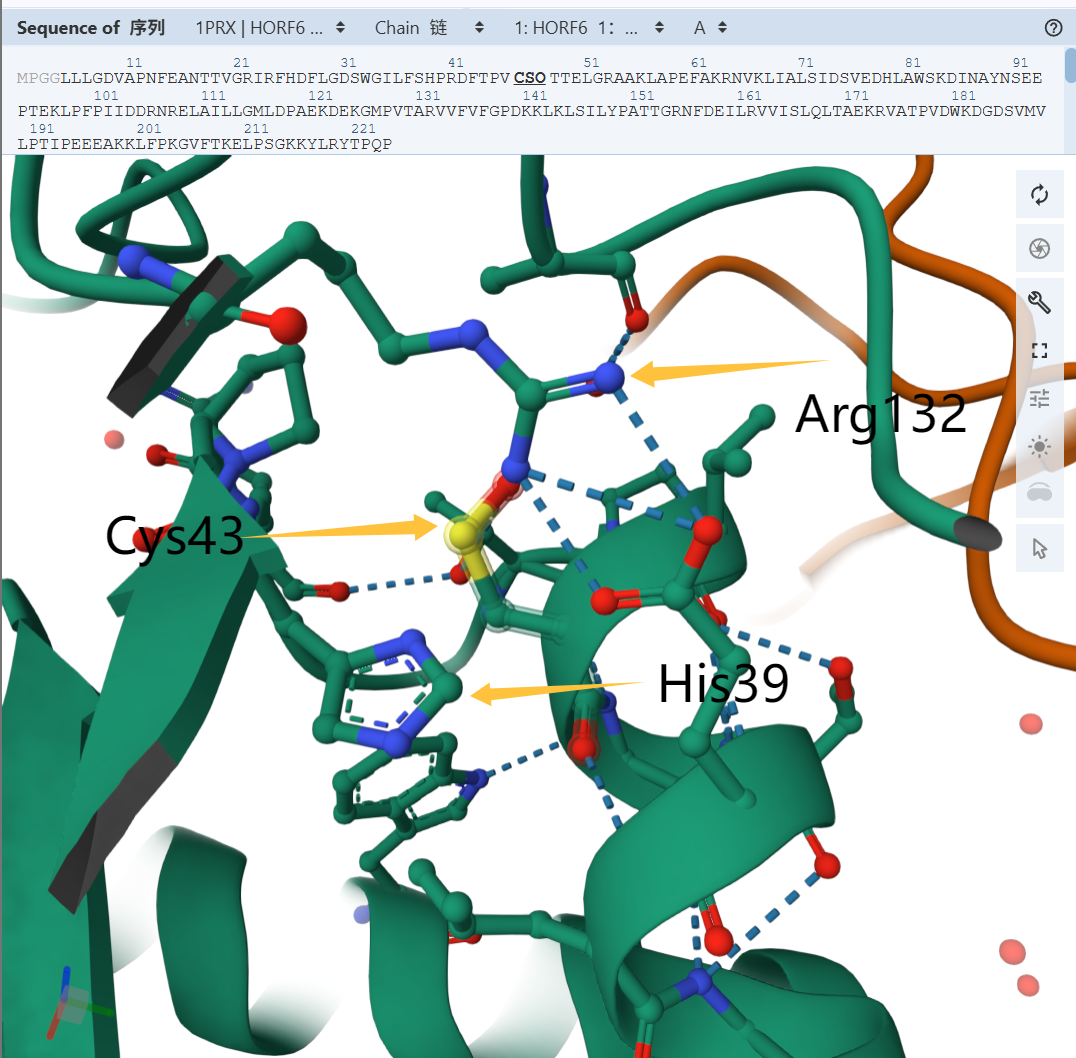
遇事不决,蛋白结构学。PrxVI 的晶体结构解析显示,\(\ce{C_P}\) 的硫原子被两个碱性氨基酸残基 \(\ce{His^{39}}\) 与 \(\ce{Arg^{132}}\) 包围,位于一个狭窄口袋的底部,碱性氨基酸残基带来的 正电荷 环境稳定了硫醇盐阴离子,降低了 \(\ce{pK_a}\),促进对过氧化物亲核进攻和 \(\ce{C_p-SOH}\)的生成。
Prx 活性中心
\(\ce{His^{39}}\) 的咪唑环作为质子受体接受 \(\ce{C_P-SH}\) 解离出的 \(\ce{H+}\),\(\ce{Arg^{132}}\) 的胍基带有正电荷,稳定了 \(\ce{C_P-S-}\),使得 \(\ce{C_P-SH}\) 的 \(\ce{pK_a}\) 下降。
硫醇盐阴离子 \(\ce{-S^-}\) 具有更强亲核性,反应活性更强。
当然,以上结构分析只能解释 Prx 如何选择性地将特定 Cys 残基作为活性中心,但不能解释其对 \(\ce{H2O2}\) 等过氧化物有如此之高的亲和力。
对此,有研究发现,所有 Prx 都有一个跨物种保守的活性中心序列: PXXXTXXC,以及一个距离此序列较远的 Arg,在折叠后位于保守序列附近。Prx 借此在口袋中心形成了如下 氢键网络:
- \(\ce{C_P}\) 硫醇阴离子
- 过氧化物 (ROOH)
- PXXXTXXC 中的 Thr 与 Pro
- 保守的 Arg 残基
这样的氢键网络给了过氧化物一个“着陆点”,让过氧化物可以顺利地结合到口袋中心,并接受硫醇的进攻。

第二个 Arg
2-Cys Prx 家族成员与 1-Cys Prx 家族含有第二个保守的 Arg 残基,一般认为在第一个保守 Arg 残基下游 23 个氨基酸残基。
推测该精氨酸可以向过渡态中间体提供另一个氢键。
4. Reversible Hyperoxidation of the Peroxidatic Cysteine of 2-Cys Prx
我们知道,\(\ce{C_P-SH}\) 先被氧化为 \(\ce{C_P-SOH}\),不过当二硫键形成足够慢时,\(\ce{C_P-SOH}\) 会与过氧化物进一步反应形成 \(\ce{C_P-SO2H}\) 形式的过氧化 Cys 残基。这导致在研究 Prx 动力学中,研究者们发现其过氧化物酶活性随时间逐渐降低。
为了详细讲解这个现象,让我们以 2-Cys Prx 为例。我们规定 Prx 具有 完全折叠 (FF) 与 局部展开 (LU) 两种构象状态。FF 构象是过氧化物结合口袋形成所必需的,此时 \(\ce{C_P-SH}\) 被氧化为 \(\ce{C_P-SOH}\) 。而 \(\ce{C_P-SOH}\) 要与 \(\ce{C_R-SH}\) 反应(可能在同一亚基/另一亚基/其他蛋白上),则必需从 FF 构象转换为 LU 构象,从而离开活性位点口袋。在二硫键形成之前,两个构象动态平衡,二硫键被 Trx 还原后,FF 构象重新建立。

尽管在不同 Prx 之间 LU 构象存在些许的不同,但一致在其性位点口袋不再形成,以及 PXXXTXXXC 片段重排以暴露 \(\ce{C_P-SOH}\) 侧链。我们知道,FF 构象具有过氧化物的结合位点,而 \(\ce{C_P-SH}\) 初始氧化速率十分之快,倘若当第一次氧化发生后,构象未能及时转变为 LU 构象,就会给第二个过氧化物可乘之机,导致过氧化的发生。
在真核生物中,2-Cys Prx 存在两个阻碍序列(GGLG modify 与 C 端附近的 YF modify),使得局部展开缓慢,致使过氧化相比不具有这些阻碍结构的原核生物更容易发生。
秋裤套棉裤,必定有缘故。真核生物对过度氧化的敏感性是进化过程中选择的一种特征。除了以上两个序列以外,Prx 存在更多的其他结构来调节过度氧化反应。在人中,PrxII 比 PrxIII 更容易发生过氧化。
拯救过氧化 Prx
一般情况下,二硫键的还原主要依赖于 Trx 出手,但面对出于过氧化态的 Prx,就需要更专业的 硫氧还蛋白还原酶 (Srx),其会消耗一分子 ATP ,γ-磷酸基团用于活化磺酸基,后借助 GSH 或 Trx 的还原当量将 Prx 从过氧化态中“拯救”出来。
如上所述,Prx 的过氧化本身是一个 可逆失活 的过程。对此有人提出,真核生物以此机制,使得可以在特定条件下积累 \(\ce{H2O2}\) 到相当高的水平进行信号传导。同时 2-Cys Prx 二聚体与十聚体形式也取决于 \(\ce{C_P}\) 的状态 (通常氧化态有利于二聚体,还原态有利于十聚体),推测这种过度氧化态可能影响到二聚体或十聚体的形成,从而影响 2-Cys Prx 可能的分子伴侣蛋白作用。
既往实验中,正常饲养的 Srx KO 小鼠的组织中并未如预期般大量的 \(\ce{PrxI-SO2H}\) 与 \(\ce{PrxII-SO2H}\)。但在正常生理状态下,如红细胞、线粒体部位,可以观察到 PrxII/PrxII 的过氧化态,并存在昼夜节律的特点。主要在内质网中分布的 PrxIV 在缺乏 Srx 的情况下也未观察到过氧化态。这些发现揭示了 过氧化态并非纯粹的病理性副产物,而可能是一种关键的调控机制。
5. Circadian Oscillatiohn of Hyperoxidized PrxII in Red Blood Cells
在一开始,人们都认为生物体的昼夜节律是由转录-翻译反馈环路驱动的,导致时钟基因的节律性表达。但研究者一样发现了人类红细胞中存在 \(\ce{Prx-SO2H}\) 丰度的节律性变化,这表明 Prx 过度氧化循环代表了一种保守的、祖先的昼夜节律机制。
但问题是究竟是哪种亚型起主要作用?振荡的意义又是什么?
研究者在小鼠红细胞中研究振荡机制,血红蛋白是最主要的 \(\ce{H2O2}\) 来源。而在红细胞中,主要由过氧化氢酶、GPx1 和 Prx 负责清除 \(\ce{H2O2}\)。在此三者中,具有 \(\ce{H2O2}\) 高亲和力位点的 Prx 在低浓度下更加高效。在红细胞中表达的三种 Prx 蛋白(PrxI、 PrxII 和 PrxVI)中,经证明 PrxII 是核心的振荡分子。
Srx KO 的红细胞也存在 \(\ce{PrxII-SO2H}\) 的昼夜节律振荡,并且不影响振幅与振荡的衰减速率(即使基础水平提高了),这与 Srx 依赖性昼夜节律振荡(小鼠肾上腺、心脏与棕色脂肪组织)形成了鲜明对比。
20S 蛋白酶体负责 \(\ce{PrxII-SO2H}\) 振荡的衰减阶段。这种对 \(\ce{PrxII-SO2H}\) 的选择性降解与以下事实相符:20S 蛋白酶体识别因蛋白质轻度氧化而暴露的疏水残基,以及 Prx 的过度氧化会暴露驱动寡聚化的疏水区域。

在红细胞中,每日大约 1% 的 PrxII 参与到振荡中,由于 S20 蛋白酶的降解,这种振荡周期会导致 PrxII 在红细胞的生命周期里逐渐减少,就像一个精密计时的钟表一样。此外,鉴于 \(\ce{PrxII-SO2H}\) 能特异性地与蛋白质二硫键异构酶 ERp46 结合,可以推测 \(\ce{PrxII-SO2H}\) 可能与 ERp46 等靶蛋白结合,并通过其分子伴侣活性保护邻近蛋白免受氧化变性。
Circadian Oscillation of Hyperoxidized PrxIII in Adrenal Gland, Heart, and Brown Adipose Tissues
研究者通过 WB 分析了正常生理状况下的小鼠各种组织中是否存在高氧化态的 2-Cys Prx,仅发现了对应线粒体 PrxIII 高氧化态条带,且在肾上腺中强度较高。推测是在皮质类固醇激素合成的过程中,以 CYP 存在的电子泄露为主要源头产生的 \(\ce{H2O2}\) 优先被含量丰富的 PrxIII 清除。当 \(\ce{H2O2}\) 生成速率过高,致使 PrxIII 发生过度氧化失活,导致 \(\ce{H2O2}\) 溢出线粒体至胞质中。\(\ce{H2O2}\) 作为信号分子激活上游的凋亡信号调节激酶 1 (ASK1),磷酸化激活 p38 MAPK,从而激活 p38,抑制类固醇生成急性调节蛋白 (StAR) 的活性,导致皮质酮的产生下调。
在此上过程中,ASK1 还会抑制一种双特异性磷酸酶来延长 p38 磷酸化状态的持续时间。
皮质类固醇激素的合成
肾上腺皮质的丝状带与束状带负责将 胆固醇连续氧化为皮质类固醇(糖皮质激素/盐皮质激素)。该过程需依赖特定的细胞色素P450 (CYP) 单加氧酶和脱氢酶,通过碳链裂解、羟基化及脱氢等连续氧化反应来实现。
- 胆固醇向孕烯醇酮的转化(起点与限速步骤):胆固醇转运至线粒体后,由胆固醇侧链裂解酶(CYP11A1 / P450scc)催化,连续进行两次羟基化并裂解侧链,切除6个碳原子,生成含21个碳原子的孕烯醇酮(Pregnenolone)。类固醇合成快速调节蛋白 (StAR)介导的胆固醇线粒体转运途径是核心限速步骤。
- 孕烯醇酮向孕酮的转化(共同通路):孕烯醇酮在内质网的3β-羟基类固醇脱氢酶(3β-HSD)与异构酶的协同作用下,被氧化脱氢生成含21个碳的孕酮(Progesterone),这是所有皮质类固醇合成的共同前体。
- 盐皮质激素合成:
- 第一步(羟基化):在球状带中,孕酮在C21位由21-羟化酶(CYP21A2)氧化,生成11-脱氧皮质酮(DOC)。
- 第二步(再次羟基化):11-脱氧皮质酮(DOC)在C11位由11β-羟化酶(CYP11B1)氧化,生成皮质酮。
- 第三步(最终氧化):皮质酮在C18位由醛固酮合酶(CYP11B2)进行氧化与甲基羟基化,最终生成醛固酮(Aldosterone)。
- 糖皮质激素:
- 第一步(C17羟基化):在束状带中,孕烯醇酮在C17位由17α-羟化酶(CYP17A1)氧化生成17α-羟孕烯醇酮,随后经3β-HSD转化为17α-羟孕酮。
- 第二步(C21羟基化):17α-羟孕酮由21-羟化酶(CYP21A2)氧化生成11-脱氧皮质醇。
- 第三步(C11羟基化):11-脱氧皮质醇由11β-羟化酶(CYP11B1)氧化,最终生成皮质醇(Cortisol)。
促肾上腺皮质激素 (ACTH) 和皮质醇浓度由下丘脑-垂体-肾上腺 (HPA) 轴驱动昼夜节律变化,并受下丘脑视交叉上核 (SCN) 的主生物钟控制。小鼠肾上腺中 \(\ce{PrxII-SO2H}\) 和磷酸化 p38 的丰度也呈现昼夜节律性波动,而 p38 和 PrxIII 的总量保持不变。类固醇生成细胞特异性敲除 Srx 可阻止 \(\ce{PrxII-SO2H}\) 和皮质酮水平的昼夜节律性变化。以此证明 PrxIII 的可逆性过度氧化通过 \(\ce{H2O2}\)/p38/StAR 通路为类固醇生成的昼夜节律振荡提供了一种 负反馈调节机制。
研究者同样在棕色脂肪组织、心脏和肺组织中的线粒体组分检测到了 \(\ce{PrxII-SO2H}\) (虽然含量极低)丰度的日波动,并同样观察到了线粒体中 Srx 含量的日波动,且与前者呈 反相。这些观察结果表明,线粒体中 Srx 的丰度是 PrxIII 高氧化状态的关键决定因素,并因此决定了线粒体 \(\ce{H2O2}\) 的释放时间与程度。
与肾上腺皮质类似,棕色脂肪组织与心脏高水平的脂肪酸代谢,伴随有呼吸链电子泄露,导致 \(\ce{H2O2}\) 的生成。在中枢与局部外周生物种调控之外,生理条件下的 PrxIII 和 Srx 同样控制着它们线粒体向外释放 \(\ce{H2O2}\) 的时间和程度,这将胆固醇代谢与能量代谢同生物钟联系起来。
线粒体 Srx 日波动
Srx 是一种胞质蛋白,当氧化应激发生时,\(\ce{H2O2}\) 促进 Srx 与 HSP90 形成二硫键,通过 TOM 被转运到线粒体中。HSP90 的辅助伴侣蛋白 FK506 也参与了此过程。被转运入线粒体的 Srx 将 \(\ce{PrxII-SO2H}\) 还原,随着后者的减少,Srx 更容易被蛋白酶 Lon 降解,从而开始下一轮振荡循环。
7. Intracellular Messenger Function of Hydrogen Peroxide and the Role of Peroxiredoxins
\(\ce{H2O2}\) 作为胞内信使,与其他信号分子不同的是,请并非直接与效应分子结合,而是通过氧化对方的关键残基——比如 Cys,来发挥调控作用的。但是对于大多数靶蛋白的目标 Cys 残基来说,其与 \(\ce{H2O2}\) 的反应活性远低于 Prx 的 \(\ce{C_P-SH}\)。这意味着 Prx 有时候需要暂时“退出舞台”(也就是失活),来让 \(\ce{H2O2}\) 的浓度升高到一定水平,并由足够的时间作用于靶蛋白上。对此细胞存在有至少两种经典路径:
- 质膜上肉瘤家族蛋白酪氨酸激酶的磷酸化
- 细胞周期蛋白B (CycB) 依赖性激酶的磷酸化
还有另一种情况,就是 \(\ce{H2O2}\) 并不直接氧化靶蛋白残基,而是通过 Prx “借力打力”。\(\ce{H2O2}\) 氧化 Prx,再由 Prx 将氧化状态转移给靶蛋白。这样做的好处是 Prx 对 \(\ce{H2O2}\) 极其敏感,使得这种信号传导途径在低浓度下也有效。在细胞中经典的有 ASK1 与 STAT3 两种蛋白受到此种调控。
8. Localized Regulation of Hydrogen Peroxide Concentration Around Lipid Rafts Via Tyrosine Phosphorylation of PrxI
许多哺乳动物细胞在刺激下会产生 \(\ce{H2O2}\),而 \(\ce{H2O2}\) 可作为信号分子发挥作用。这些依赖于受体刺激产生的 \(\ce{H2O2}\) 产生于 质膜脂筏上的NADPH氧化酶复合物。一个典型的案例是生长因子刺激的细胞中受体信号的传播依赖于 \(\ce{H2O2}\) 的产生与积累,源自于被激活的受体酪氨酸激酶 (RPTK) 不足以提升胞质中靶蛋白磷酸化稳态水平,需要依赖 \(\ce{H2O2}\) 氧化蛋白酪氨酸磷酸酶 (PTP) 的 Cys 来抑制其活性。
研究者发现,细胞中仅占 0.3% 的 \(\ce{Tyr^{194}}\) 磷酸化 PrxI 聚集于脂筏附近,表明 PrxI 的失活对 \(\ce{H2O2}\) 积累的关键作用。同时被激活的 RTPK 也会激活 Src 家族激酶,诱导 PrxI 磷酸化,而 \(\ce{H2O2}\) 的积累也会激活 Src 家族激酶,形成了正反馈通路。

9. Localized Regulation of Hydrogen Peroxide Concentration Around Lipid Rafts Via Cdk1-Dependent Phosphorylation of PrxI
哺乳动物的 2-Cys Prx 包含一个可被 Cdks 磷酸化的共有序列。在有丝分裂早期,Cdk1 特异性磷酸化并灭活了定位于中心体处的 PrxI(Thr90 位点),从而允许局部积累的 \(\ce{H2O2}\) 进行局域性信号调控,并在有丝分裂晚期通过去磷酸化恢复对中心体的抗氧化保护。
同时 \(\ce{H2O2}\) 在细胞周期中发挥重要的作用,在有丝分裂初期,Cdk1 磷酸化抑制 PrxI 使得 \(\ce{H2O2}\) 积累,从而使得 Cdc14B 的失活,并使得中心体上 APC 和 Cdh1 结合被阻断。而在有丝分裂后期,PrxI 再次被激活,以避免中心体暴露在高浓度 \(\ce{H2O2}\) 下。
10. Peroxiredoxins As Sensors and Transducers of Signaling by Hydrogen Peroxide
除开局部失活以保护 \(\ce{H2O2}\) 以外,Prx 还可以通过被 \(\ce{H2O2}\) 氧化后,将氧化态传递给靶蛋白的方式传递信息。哺乳动物中第一个被发现具有如此感受器作用的是 PrxIV,主要定位于内质网腔。分泌蛋白和膜蛋白均在内质网中合成并经历氧化蛋白折叠,后者涉及将电子从未折叠蛋白的还原型半胱氨酸残基转移到蛋白二硫键异构酶 (PDI) 的分子内二硫键上,而后者二硫键的重新形成依赖于将电子转移到 \(\ce{O2}\) 上,该过程由内质网膜相关氧化还原酶1 (Ero1) 催化,生成 \(\ce{H2O2}\),其可以被 PrxIV 清除。
但在内质网中并不含有 Trx,这意味着氧化态下的 PrxIV 无法被还原,但 PrxIV 有办法,它会和 PDI 做一个等价交换,将自身的氧化态传递给还原态的 PDI,从而实现双赢。

PrxI 参与到 ASK1 间的二硫键形成,介导脱嘌呤/脱嘧啶核酸内切酶 1/氧化还原因子 1 (APE1/Ref-1)的氧化。通过瞬时形成二硫键连接的 PrxII-STAT3 中间体催化 STAT3 分子间二硫键形成,产生不具有转录激活活性的 STAT3 二聚体。
许多蛋白质的功能受半胱氨酸残基可逆氧化的调控。仅就蛋白激酶而言,IκB 激酶α/β 、Src 家族成员、ATM(共济失调毛细血管扩张症突变蛋白)以及受体酪氨酸激酶 RET(转染重排蛋白)已被证实可通过 H₂O₂ 介导的半胱氨酸氧化而被激活。